Orecchio interno
A questo livello si verifica la conversione dell’onda acustica in stimolo elettrochimico che può essere trasmesso al sistema nervoso centrale. Durante tale processo , l’orecchio interno analizza lo stimolo sonoro in termini di frequenza, intensità e proprietà temporali.
La coclea presenta tre condotti riempiti di liquido, funzionalmente distinti in :
-uno spazio perlinfatico, comprendente la scala timpanica e la scala vestibolare in comunicazione tra di loro e contenenti perilinfa.
-uno spazio endolinfatico ( scala cocleare), isolato dai precedenti e contenete endolinfa.
La differenza di composizione elettrolitica crea un ambiente elettrochimico che rende possibile la trasudazione neurosensoriale.
L’endolinfa ha una composizione elettrolitica simile a quella dei fluidi intracellulare ( ricca in potassio e povere in Sodio e Calcio), mentre la perilinfa l’ha simile a quella dei fluidi extra-cellulari (ricca in sodio e povera in potassio).
Tale composizione subisce solo lievi variazioni a livello delle due scale perilinfatiche, così come tra il giro basale e l’apicale della rampa cocleare. Tale omeostasi è garantita attraverso un flusso radiale nei differenti punti labirintici, a livello locale basandosi sul fatto che:
-esiste una barriera anatomica tra endolinfa e perilinfa (membrana di Reissner, stria vascolare e lamina reticolare formata da giunzioni intercellulari serrate).
-nonostante tale barriera, gli elettroliti subiscono una lenta perdita, dando luogo a gradienti di concentrazione che si estrinsecano in una corrente continua registrabile all’interno della coclea.
– gli elettroliti che fluiscono nella perilinfa ritornano all’endolinfa attraverso il legamento spirale e la stria vascolare, ove risiedono un sistema enzimatico ed organuli cellulari necessari al mantenimento dell’omeostasi. Dal momento che il passaggio di potassio nell’endolinfa- contro un gradiente di concentrazione- richiede energia, è fondamentale l’azione dell’enzima Na/K/ATPasi ( localizzato nelle cellule marginali della stria vascolare e nel sottostante legamento spirale) che utilizza allo scopo i depositi energetici prodotti dai mitocondri della stria e del legamento spirale.
Le differenze in contenuto elettrolitico delle diverse scale labirintiche danno luogo, come detto , a gradienti di concentrazione tra gli spazi endolinfatico e perilinfatico. Dal momento che gli elettroliti sono particelle cariche (ioni), tali differenze creano altresì dei potenziali elettrochimici tra perilinfa ed endolinfa. Von Bekesy ha in proposito identificato nella scala vestibolare un potenziale di +5 mV rispetto alla scala timpanica, mentre nella scala media (cocleare) è stato registrato un potenziale positivo relativamente ampio, pari a + 80 mV, corrispondente al potenziale endococleare che serve come forza trainante per la trasduzione del segnale.
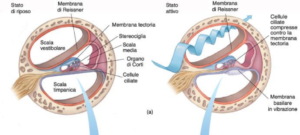
Come detto, attraverso la finestra ovale l’onda sonora viene trasmessa alla scala vestibolare, generando una serie di onde nella perilinfa che causano un depiazzamento della membrana basilare e dell’organo di Corti e che in ultima analisi, causano l’estroflessione nell’orecchio medio della membrana della finestra rotonda ( timpano secondario di Scarpa). La frequenza di movimento della membrana basilare è in relazione alla frequenza dello stimolo sonoro e rappresenta il primo processo di trasduzione. Secondo la teoria di Bekesy (1960), l’ampiezza dell’onda viaggiante lungo la membrana basilare aumenta fino a raggiungere un massimo specifico per frequenza (frequenze acute alla base, frequenze gravi all’apice della coclea), per poi declinare (proprietà di sintonia passiva). Studi successivi hanno dimostrato che l’ampiezza dell’onda viaggiante non aumenta gradualmente ma, dopo aver viaggiato lungo la membrana basilare determinando un minimo depiazzamento, raggiunge improvvisamente un picco massimo nel punto della membrana basilare sensibile per frequenza a quella dello stimolo e poi declina successivamente in modo altrettanto rapido.
Quindi la membrana basilare agisce come un filtro passa-banda finemente sintonizzato: siti specifici corrispondono a frequenze specifiche.
Questo meccanismo di fine sintonia dipende da processi attivi, che richiedono energia (che, dunque, non potevano essere riscontrati negli studi su cadavere di von Bekesy), al livello delle cellule ciliate esterne. La superficie apicale delle cellule ciliate, con le loro stereociglia, è bagnata nell’endolinfa ed è dunque esposta al potenziale endococleare (+80mV) mentre al loro interno il potenziale di riposo è di circa -40,-60 mV: la differenza di potenziale attraverso la membrana apicale delle cellule ciliate è quindi pari a 120-140 mV.
Da queste misurazioni prende avvio la teoria di Davis (1958) , secondo la quale la stria vascolare rappresenta la maggiore fonte d’energia per il mantenimento positivo endococleare. La superficie apicale della cellula ciliata funziona come resistore variabile, la cui impedenza è variata dal depiazzamento meccanico delle stereociglia. Poiché i corpi delle cellule ciliare si muovono con la membrana basilare sotto lo stimolo acustico, mentre le stereociglia si trovano in ambiente statico, circondate da endolinfa e nella membrana tectoria, la vibrazione della membrana basilare determina un movimento delle cellule ciliate rispetto alla membrana tectoria dell’endolinfa.
-Se la deflessione delle stereociglia avviene verso la pila più alta, si determina l’apertura dei canali che facilitano l’ingresso di K, grazie ad un gradiente di 120-140 mV , con depolarizzazione delle cellule ciliate.
– Se la deflessione si verifica verso la direzione opposta , si crea un’iperpolarizzazione per la chiusura di questi canali perennemente aperti , anche in condizioni di riposo, impedendo così ulteriormente il flusso di K.
Dunque l’apertura e la chiusura dei canali del potassio modulano le variazioni di potenziale elettrochimico delle cellule ciliate attraverso un processo meccanico.
Quando le stereociglia si piegano verso la fila più alta, le “tip links”, una sorta di molle situate tra il centro di ciascun stereociglio e l’apice di uno adiacente, più basso , si contraggono ed i canali vengono mantenuti aperti.
Quando si piegano dalla parte opposta , le “tip links” si rilasciano ed i canali si chiudono.
La depolarizzazione determina effetti differenti sui due tipo di cellule ciliate.
Le CC interne funzionano come recettori sensoriali e quindi la loro depolarizzazione dà luogo ad un’attivazione delle fibre nervose afferenti con trasmissione del segnale uditivo al SNC;
-le CC esterne hanno invece una minima funzione sensoriale , regolando le proprietà motorie responsabili per una fine sintonia della coclea.
La stimolazione di una cellula ciliata dà luogo ad un potenziale di recettore cellulare che ha 3 compenenti:
- Una risposta fondamentale , rappresentata dalla depolarizzaizone, la cui ampiezza dipende direttamente dell’intensità dello stimolo fino ad un pinto di saturazione, passato il quale non si hanno ulteriori variazioni.
- Un potenziale a corrente alternata che si correla con la frequenza dell’onda sonora stimolante , come cicli di depolarizzazione-polarizzazione.
- Una variazione DC del potenziale delle cellule ciliate, che possono depolarizzarsi o iperpolarizzarsi secondo la direzione della deflessione delle stereociglia, con ampiezza di depolarizzazione maggiore di quella dell’iperpolarizzazione, ad ogni gradi di deflessione.
- La depolarizzazione delle cellule ciliate interne dà luogo all’attivazione dei canali ionici lungo la parete cellulare laterale. Questi canali permettono l’uscita di K e l’ingresso di calcio nella cellula con rilascio di neurotrasmettitori (glutammato) della base cellulare, in proporzione all’intensità dello stimolo. Il neurotrasmettitore si lega alle terminazioni nervose afferenti alla base cellulare e dà luogo ad un potenziale d’azione propagato alle fibre afferenti. La depolarizzazione delle cellule ciliate esterne segue un processo analogo, ma con risultato differente. Il loro ruolo principale è infatti quello di convertire il segnale acustico in elettrochimico , trasmissibile a SNC. La maggior parte delle fibre afferenti (95%) contrae sinapsi con le CC interne e seve per trasmettere il segnale sensoriale. Al contrario le CC esterne hanno minimi contatti con le fibre afferenti e sono quasi esclusivamente collegate con le fibre efferenti. Il ruolo delle CC esterne è quello di dare alla coclea le proprietà di fine sintonia, permettendo a ciascuna regione cocleare di corrispondere ad una frequenza specifica. Questa proprietà di amplificazione è ottenuta dalle proprietà di allungamento e contrazione derivanti dai processi di depolarizzazione ed iperpolarizzazione, che aumentano il depiazzamento della membrana basilare.
Fonte Dott. Maurizio Bambara.